
僕が飛ばしているセスナ172の燃料タンクは翼の中にあります。
高翼機ですから、翼の上にある燃料キャップから給油するためには機体によじ登らなければなりません。その足がかりとして、翼を支える梁(ストラットと呼びます)の真ん中あたりに、ラバー素材っぽいステップがあります。

これが意外な使い道で役に立つのです。
投稿者「始祖鳥」のアーカイブ
丘珠急行
ヤポンスキー(叔母ともいう)に一泡吹かせるために、11月10日の10時過ぎくらいに西野第二小学校上空で示威飛行をしてきたスキー(・∀・)

MSFSで倉庫潜りをお見せしますよ!(・∀・)
前回、前々回はActualの飛行でしたが、今度はシミュレータでの飛びをお見せします。
Extra300Lでパターンを変えて4回、倉庫潜りをやってみました。(・∀・)
twitvideoで見る
明日はNZの野鳥の動画をアップしようと思っています(・∀・)
NZでの僕の飛びっぷりを見るが(・∀・)イイ
NZでのかつての僕の仕事の一部をお見せします。飛行中に他の飛行機を追い抜く映像です。僕はこのとき機長席(左席)でカメラ回してます(・∀・)
twitvideoで見る>
場所は南島西海岸のハーストからワナカ方面に移動する途中、マウント・アスパイアリング付近です。
雨竜沼湿原上空、10月21日
10月21日に雨竜沼湿原の上空を飛んだときの動画です!カメラマン兼パイロットです(・∀・)
twitvideoで見る
もう紅葉する樹は全て葉っぱを落とし、常緑樹だけが残っているのでこんな風に見えます。
メモ用のブログを作ってみました
twitterやTINAMIから来る場合のクッションURLとして、このブログを作ってみました。うまく設定できたかな?(・∀・)
リムサウルス(Limusaurus)の退化した親指――鳥類の恐竜起源仮説に新たな証拠
「Nature」6月18日号で、リムサウルス・イネクストリカビリス(Limusaurus inextricabilis)という新種のケラトサウルス類が発表されました。
属名のLimusaurusは「泥(にはまった)トカゲ」、種小名のinextricabilisは「逃げられなかった」の意味で、湿地の穴にはまってそのまま化石化したと見られる産状に由来しています。中国北西部、新彊の1億5900万年前の堆積層から発見されたこの恐竜は歯を持たず、そのかわりに嘴を持っています。発表者の徐星(Xing Xu)は、鋭い鉤爪を持たない腕、砂嚢石(gizzard stones)の存在から、この恐竜が獣脚類としては珍しい植物食であったと考えているようです。
「草食系」の獣脚類という時点で既にユニークなのですが、これに加えてリムサウルス(Limusaurus)には進化史の上で重要な特徴を持つという一面があります。退化の過程にある第1指(親指)の痕跡が残っていたのです。
これが一体何を意味するのかについて、以下で説明します。
現在では「恐竜の子孫である」という仮説が共通認識であり、「鳥類は恐竜の子孫ではなく、恐竜以外の樹上性の主竜類に起源を持つ」と主張する研究者も今では殆ど残っていませんが、アラン・フェドゥーシア(Alan Feduccia)やラリー・マーティンなど、メディア露出が多く声の大きな研究者がこの説を主張しているため、この仮説がメディアから完全に姿を消すことは当分はなさそうです。
「恐竜は鳥の祖先ではない」――その根拠として大きなものの一つが前肢の指の同一性についてのものでした。彼らの意見をおおむね要約すると下記のようになります。
鳥類の祖先とされる非鳥類型獣脚類(恐竜)の3本指は、第1,2,3指(親指、人差し指、中指)であると同定されている。
これに対して、鳥類の胚の発生過程で翼に残るのは第2,3,4指(人差し指、中指、薬指)である。
ゆえに鳥類の祖先は獣脚類(恐竜)ではない。
恐竜の指が第1,2,3指(親指、人差し指、中指)で構成されているという主張の根拠は、図版(Feduccia,2002)を見ると理解しやすいです。
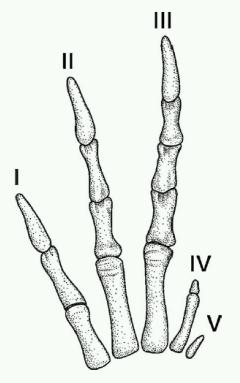

青い画像は鳥類の胚の発生過程で、第2,3,4指(人差し指、中指、薬指)が残っていることが分かります。
ところが、手先の骨格図は三畳紀後期の恐竜、ヘレラサウルス(Herrerasaurs)のものですが、第4指(薬指)はかなり退化傾向にあり、第5指(小指)は退化寸前まで縮小しています。この事実は、恐竜が第4,5指(薬指、小指)を失う途中経過を示している、というのが彼らの主張です。(Natureのトピックスで言及されているのはジュラ紀初期の恐竜、ディロフォサウルス(Dilophosaurus)ですが、この種にも同様の特徴があるようです。)
この疑義に対する「恐竜は鳥の祖先」支持者からの反論は複数件あり、その論旨はおおむね2種類に分類できます。
(1)発生過程は必ずしも進化の過程を反映するわけではない。
(2)実は恐竜の指も鳥類と同じ第2,3,4指(人差し指、中指、薬指)である。
ここで記事の初めに書いたことに戻りますが、今回発表されたリムサウルス(Limusaurus)には、「退化の過程にある第1指(親指)の痕跡が残っていた」――つまり今回の発表は上記(2)に該当するものです。基盤的な獣脚類と鳥類を結ぶミッシングリンクということになります。
今回の徐星(Xing Xu)の主張が正しいとすると、考えられる影響は下記の2点です。
第一に、「鳥類の祖先は恐竜である」という仮説の支持がさらに強まり、「恐竜ではない」派は仮設を支持する有力な証拠を失うことになります。
第二に、獣脚類の指の消失過程が下記(1)(2)のどちらだったかについて、新たな疑問が生まれることになります。
(1)鳥類に至る獣脚類とそれ以外の獣脚類は共に3本指であるが、それぞれ指の減らし方が違っていた
(2)ヘレラサウルスを含む獣脚類の共通の系統において、第4指の退化傾向が逆転し、第1,5指が失われた
今後の議論が興味深いニュースでした。
【参考文献】
1. Xu, X. et al. Nature 459, 940-944 (2009).
2. Feduccia, A. et al. “The hand of birds revealed by early ostrich embryos”, Naturwissenschaften 89, 391–393 (2002)
【参考URL】
▼Dinosaur’s digits show how birds got wings (Natureのハイライト)
▼<恐竜>「鳥の指」持つ新種、進化の証拠を補強 中国で発見(6月18日9時6分配信 毎日新聞)
▼鳥と同じ「指」持つ恐竜=進化の物的証拠に-米中チーム(6月18日5時37分配信 時事通信)
▼ニュース – 古代の世界 – 新しい恐竜化石が示す鳥類の祖先(記事全文) – ナショナルジオグラフィック 公式日本語サイト
「新発見は恐竜-鳥類の繋がりに新たな疑いを提起する」のかなあ?
ScienceDailyを読んでいたところ、『新発見は恐竜-鳥類の繋がりに新たな疑いを提起する』とのコラムに出会いました。この記事を読んでも明確な理由が書いてあるわけではなく、どうにも要領を得ません。
こういう場合は元の論文を確かめるのが早道というわけで、アブストラクトを軽く翻訳してみました。
ワニ類の肺と心臓血管の器官は,鳥類の心臓と肺気嚢システムほどには著しく専門化していないが,現生の全ての主竜類は4室の心臓と,異なった起源を持つ血管の張り巡らされた蜂の巣状の肺を持っている.鳥類において,通常の肺機能は,背面に位置した毛細血管の分布が行き渡らない腹部の気嚢が,発達した胸骨および特別にヒンジの付いた肋骨によって換気されることを要求する.薄い壁で囲われ,容積のある腹部の気嚢は,負の(吸入側の)圧力の発生の間,内部の崩壊を妨ぐために,横方向と尾方向に支えられる:背部に方向付けられた,横に開いている下腹部である複合仙骨,そして特殊化した大腿-腿複合体は,必要な支持を提供し,主として吸入崩壊を防止する.比較の結果,おそらく獣脚類恐竜は,同様に拡大された腹部の気嚢と,彼らの呼吸と整合性がある骨-筋肉の変異に欠けていた.拡張された,機能的な腹部の気嚢がないため,獣脚類が鳥類的な気嚢肺を所有する可能性は少なかった.獣脚類における鳥類的な肺の機能の欠如の可能性は,いっそう洗練された心臓血管の解剖学的特徴の示唆と一致しない.
http://www3.interscience.wiley.com/journal/122395783/abstract?CRETRY=1&SRETRY=0
(筆者による訳出)
筆頭者はQuick博士、共同著者のRuben教授は鳥類の心肺関係の研究で頻繁に名前を目にする研究者ですね。
さて、これがなぜ『この発見はおそらく、鳥類が恐竜と並行の経路で――多くの恐竜種が存在する前にそのプロセスを開始して――進化したことを意味する。』(This discovery probably means that birds evolved on a parallel path alongside dinosaurs, starting that process before most dinosaur species even existed.)となるのかが不明です。
尾が短くなったことによる重心位置調整のために、大腿骨を常に曲げた状態にし、膝の部分を支点にする姿勢となったと考えるのが一般的な解釈と思います。その過程で「二次的に」大腿骨による腹部気嚢の構造保持も行なうようになったと考えれば、分岐学で支持される『恐竜→鳥類』仮説に特に矛盾はないように思えるのですが、Ruben教授の『恐竜は鳥の祖先ではない』という確信は一体どこから来ているんでしょう。
岩手県立博物館に行ってきました
今日は岩手県立博物館に行ってきました。

先日の古生物学会でクジラの初期進化に関する講演を聞いて古鯨類に興味が沸いてきました。講演中に言及されていたミズホクジラ(Herpetocetus sendaicus)やマエサワクジラ(Burtinopsis sp.)などの化石鯨類のレプリカはいずれも岩手県立博物館に展示されています。まずは現物を見に行ってみよう!
ということで、初めて伺ったのが先月の3連休。
どこがどうなっているのかしっかり記憶して帰らねば。(でも鳥類とは骨が全然違うのでどうにもならない)
メモとデジカメを持って復元骨格の周りを憔悴したツキノワグマのごとくくるくる回っていると、親切な学芸員さんが声をかけてくださいまして、シリーズ化した一般配布用の資料をいただき、その上まさかお話できると思ってなかった方(※1)と直接お話できる機会まで頂きました。
岩手県立博物館の皆様、本当にありがとうございました。
そして今日の目的は、前の訪問時のお礼というわけではありませんが、前回話題にのぼった始祖鳥骨格標本(サーモポリス標本)についての論文の写しをお渡しすることと、先日うまく撮影できず心残りだった骨質歯鳥類の一種(Pseudodontornithidae gen. et sp. indet.)の上腕骨(標本番号 IPMM 40061)の写真もしっかり撮影してくることで、結論を言えば両方とも達成できました。本当に岩手県立博物館の皆様にはお世話になりっぱなしで恐縮です。
古生物学会年次総会
閉幕しました。ヽ(・v・)ノ
東北大での年次総会は1999年以来でした。今回はほとんどの時間を古環境と素敵放散虫のB会場で過ごしました。本当に勉強になりました。
とりあえず古鳥類・真鳥類とその類縁に関して興味深かった発表を忘れないうちにメモ。
・手取層群産出の未同定の小型獣脚類化石
中足骨がアークトメタターサル(発表者の方は違う言い回しで表現されていました)であることから、おそらくドロマエオサウルス類ではないか(これ以外にもアークトメタターサルな足を持つグループはあるが、他の特徴も考え合わせるとドロマエオサウルス類である可能性が高いということなのだろう)
・アヴィミムスは従来考えられていたほど鳥類に似た特徴をもっていなさそう
新たに発見された橈骨と中手骨を見る限り鳥類的というよりはむしろ基盤的獣脚類のそれに近いという発表
・エピオルニスの脳函をCTスキャンし他の走鳥類(古顎類)と比較
臭球は大きめ、下垂体は大きめ
他の走鳥類と大きくは違わない
とりあえず生存証明&出没予定
明日から3日間、古生物学会@東北大で僕と握手! ヽ(・v・)ノ
なにこれ胡散臭い(・∀・;)
研究チームは大学生341人に、まず視力テストと称して画面に連続表示されるブロックの位置を当てさせた。このときAppleまたはIBMのロゴを本人も認識できない速さで表示した。その後、「壁を作ること以外で、レンガを使ってできることすべてを挙げよ」という課題に取り組ませた。
Appleのロゴを見せられた学生は、IBMのロゴを見せられた学生と比べ、ユニークなレンガの使い方を考案したという。また判定に当たった審査員も、Appleのロゴを見た学生たちの案の方が「より創造的」との判定を下した。
ただし学生たちはなんのロゴを見たかはまったく記憶していなかった。「自分が見たロゴを言い当てられたら100ドル」としたものの、当てられた学生は1人もいなかったという。
「Apple」のロゴを見るだけで創造性が上昇――デューク大学調査
マスコミの報道は元の論文と全く違うことを言っている場合も多いですし、元の論文を読んだわけでもないのですが、この記事を読む限り非常に胡散臭い。(サブリミナル効果自体、いくつかの実験によりその存在そのものに否定的な結果が出ている)
果たして統計上有意な結果なのか、この著者らが主張する『創造性』の定義はどういうものだったのか、元の論文を読むべきなのでしょうけど、これは読むだけ時間の無駄のような気もします。
・追記
Automatic Effects of Brand Exposure on Motivated Behavior: How Apple Makes You “Think Different”
アブストラクトを読んでも、前述のITMediaの記事以上のことは言っていないですね。
バイク関連の備忘録
◆TL1000R(トキコ6POD)は下記の機種とフロントブレーキパッドの互換性あり。(デイトナ・ゴールデンパッドの型番 47370)
| GSX-R750SPR | 94-95 |
| GSX-R750 | 96-99 |
| GSX1400 | 01-07 |
| GSX1300R | 99-06 |
| GS1200SS | 01-02 |
| バンディッド1200/S | 00-05 |
| GSX-R1100W | 93-99 |
| GSX-R1000 | 01-02 |
| バルカン1500 | 01-03 |
| ZRX1200/S/R | 01-04 |
| ZRX1200R | 05-07 |
| ZX-12R | 00-03 |
| ZRX1100/II | 97-00 |
| GPZ900R(A12-A16) | 99-03 |
| ZX-9R(B3)ヨーロッパ仕様 | 96 |
| ZX-9R | 97-01 |
| ZX-7R | 96-01 |
| ZX-6R | 98-01 |


◆フォームの矯正
コーナリング時に上体が傾きすぎ。(特にドライ路面では)腰をもっとイン側に入れ上体を立てるようフォームを矯正。

◆VTR1000SP-1/2(NISSIN 4POT)は下記の機種とフロントブレーキパッドの互換性あり。(デイトナ・ゴールデンパッドの型番 47332)
| CB1300SF | 00-02 |
| CBR954RR | |
| CBR929RR | |
| CBR900RR | 98-03 |
| CBR600RR | 03-04 |
| CBR600F | 99-01 |
| CBR600F4i | 01-03 |
| Honet600 | 98-00 |
| CBR400SF | 99-06 |

◆NSR250RLのオイルポンプの修理
部品取り寄せること。
◆スケジュール
3/29, 3/30の午前中に走行枠あり。
4/2のマイペースラップは中止。4/17にあり。
◆タイヤ交換
もう路面凍結はないと確信。CRM80のスパイクタイヤをノーマルタイヤに交換のこと。
狙い撃つぜ!(カメラで)
狙撃と撮影、英語だと両方ともshootなんですよね。
台原森林公園の地図を作る計画は着々と進行中。西側はおおまかなところをカバーできたので、今日は東側を重点的に調査しました。今回もその過程で出会った鳥たちについての覚書です。

シジュウカラです。太目の小道で6~8羽の群れが居ました。

3/9の探索時に出会ったのと同じ個体であると思います。(顔つきとか仕草とか)


樹上にダイサギも確認しました。

カルガモのつがいです。


セキレイのつがいも確認しました。

キジバトですが、前述のものとは違う個体です。餌のとり方が豪快な奴でした。鳥は個体ごとに個性があって面白いです。

距離が遠いですが、ジョウビタキの雌でした。実は当地のデスクトップキャラクター『野原ひたき』の鳥形態はジョウビタキの雌という設定です。

ヒヨドリが木に止まっていました。
台原森林公園の地図作成(3/9編)
先週の日曜日(2008年3月9日)もGPSとフィールドノートを手に台原森林公園の地図作成をやっていました。西側の小道はかなり網羅できてきたような気がします。
地図作成の過程で見かけた鳥です。

かろうじてジョウビタキと分かりますが、AFが背景の木を捉えちゃってピンぼけ。
冬も終わりだからもうしばらくしたらお別れですね。

シジュウカラの番のうちの一羽でした。

キジバトです。あまり人を恐れない不思議な子でした。
台原森林公園の地図を作成中
台原森林公園内の数箇所にある案内図は小道のディティールがそれぞれ食い違い、しかも実地を歩いた経験上どれも正しくない。
それならば正しい地図を自分で作ろうと思い立ち、ここ数週間の土曜日は登山用のハンディGPSを持って園内の水系および小道の探索を行なっています。探索の成果としての地図をhttpで公開するかは今のところ未定ですが、トラッキング情報の一部はこんな感じです。

最初は30メートル毎の移動計測設定でしたが、地図上にプロットすると座標が粗いため、今日から5秒毎の時間計測設定に切り替えてみました。計測結果は午前中だけならばメモリの容量にぎりぎり収まるようです。
地図を作る探索の過程でいろいろな野鳥と出会いました。
先週はコゲラ、カラ類(シジュウカラ、ヤマガラ)、ヒタキ(ジョウビタキの雌と思われる)、キクイタダキなどを確認しましたがカメラを持っていなかったので目視のみ。
今日はコゲラがアカマツに取り付いて摂食しているのを確認したのですが、タイミングが悪く背中しか映りませんでした。

The Evolution of Feathers
文献の流し読みをしていたところ、羽毛の起源についての面白い論文を見つけたので備忘録代わりに。
題記の論文ですが、著者はJAN DYCK、発表は1985年。22年前の論文です。
多孔性の物体はソリッドな表面よりも水をよく弾きますが、羽毛の要件はこれに適合するとのこと。
濡れた外皮から水分が蒸発した場合の体温の喪失を防ぐことができるようです。
このことから、著者は羽毛は海岸に生息する爬虫類で進化したものではないかという仮説を導いています。
大胆ですがおもしろい仮説です。
DYCK, J., The Evolution of Feathers, Zoologica Scripta 14 (2), 137-154, 1985.
トップページ改造
始祖鳥生息地のトップページをちょっと更新。
リンクにマウスカーソルが乗ったときに説明を表示する仕掛けを組み込んでみました。
http://archaeopteryx.rgr.jp/
それから地味な所で地質年代表の使い勝手を良くしてみました。層序区分毎にNAMEタグを埋め込んだので外部からの直接参照がしやすくなっています。
ZMemoスクリプト
カテゴリ作ったのに一件も記事がないので投稿。
左側の子はこねこ堂のぽちちゃん。右側がうちのひたきです。
二人とも透ける付箋紙・ZMemo上で動くスクリプトです。
ひたきはここしばらく更新をしてませんでしたが、今週末辺りに何らかの更新をしようと思っています。
中生代の古鳥類は地上性であることについての新しいアプローチ
今日はバイクでどこかに出かける予定だったのですが、天気が悪かったため、自宅でScienceDailyを流し読みしていました。その過程で興味深い論文を発見したので自分用のメモをかねての記事作成です。
Earliest Birds Acted More Like Turkeys Than Common Cuckoos(ScienceDailyの元記事)
ScienceDailyの元記事は上記URLなのですが、記事の内容が微妙にピント外れのような気がするので、論文のアブストラクトを直接訳してみました。
中生代の鳥類および非鳥類型獣脚類の採餌方法
Foraging modes of Mesozoic birds and non-avian theropods
Christopher L. Glen and Michael B. Bennett
鳥類の起源と初期進化は進化生物学上の大きなトピックである。20世紀において、進化史のシナリオでは、地表性の鳥の先祖と樹上居住性の鳥の先祖の二者が提案されていた。これは誤った2分法であると考えられる。後肢の機能を考慮に入れた場合、多くの現生鳥類が、地上と樹上の移動者というあいまいな分類にあることが問題の一部であると我々は示唆する。実際のところ、これらは互いに排他的な二者択一型の手段というわけではない。多くの現生鳥類は異なった度合いで地上~樹上の性質を示す。よって、我々は2分法よりも、現生鳥類と彼らの示す地上または樹上(あるいはその両方)の採餌行動に照らし合わせた連続体の上に配置することを提案する。この手法をテストするために、我々は完新世鳥類249種における爪先の鉤爪を分析し、樹上の採餌行動がより支配的になることに伴い鉤爪の湾曲が増加することを明確にした。 改良された鉤爪の形状測定基準は、後者の性質を表すため、現生鳥類と絶滅した鳥類との直接の比較をより多く許容する。先人の研究の対照によると、中生代の鳥類、およびそれと密接な関係のある非鳥類型獣脚類(non-avian theropod)の爪の弯曲率は、完新世の樹上性鳥類とは明らかに異なっており、それよりも「地上で採餌する(ground-foraging)」鳥類に近いことを我々は発見した。
原文(内容梗概)
(筆者による訳出:強調部分は筆者によるもの)
Current Biologyの2007年11月7日号に掲載された論文です。
爪の曲率で地上性、樹上性を推定するというアプローチは、少なくとも1984年のオストロムの論文の時点ですでに存在していたわけで、「地上で採餌したと思われる」結論も最初の論文に沿ったものなので、一体何をいまさらと思われる方もいるかもしれません。
しかし、今回の論文の新しい点は「採餌行動」を最も重要な習性と考え、「地上、樹上の主にどちらで餌をとるか」という行動様式と爪の曲率を関連付けた点です。私たちが目にすることができる現生の鳥類には、はっきり地上性とも樹上性とも言い切れないあいまいな生態のものも存在しますが、そういう鳥類もデータとして有効になるわけで、理論的には従来より精度の高い結果が得られるわけです。
その結果、どうも非鳥類型獣脚類および初期鳥類は地上で採餌する鳥類と似た傾向が見られたということですね。鳥類の初期進化を考える上で興味深い論文です。
Glen, C. L. & Bennett, M. B., 2007. Foraging modes of Mesozoic birds and non-avian theropods. Current Biology, Volume 17, Issue 21, 6 November 2007, Pages R911-R912